Evaluating PRPF4B Protein and mRNA Levels after CRISPR-Cas9 Mediated Heterozygous Knockout in Human iPSCs
Justin Nguyen
Webber Academy
Grade 11
Presentation
No video provided
Hypothesis
If one PRPF4B allele is knocked out in iPSCs, then PRPF4B mRNA and protein levels will decrease. This is because a single functional copy of PRPF4B can not produce enough stable mRNA to maintain normal protein levels.
Research
A balance of brain fluid is necessary for neurological development and function in humans.1,2 Cerebrospinal fluid (CSF) circulates within the brain’s ventricular system and performs the functions of nutrient delivery, waste removal, and the protection of neural tissues.3 Disruptions or constant changes in the regulation of CSF, whether its an overproduction, blocked fluid flow, or even a lack of fluid can create pathological fluid buildup.4 These conditions have been known to cause a range of developmental problems such as speech disorders and cognitive disorders.5 Understanding the molecular mechanisms that impact cerebrospinal fluid regulation is important for preventing these conditions and creating an effective treatment for people who are already affected.6-9
Recent research has found that RNA splicing in neurological development has been linked to CSF homeostasis.10 One gene, PRPF4B, is being investigated.11 This gene codes for a serine/threonine kinase protein that plays a role within pre-mRNA splicing.12 RNA splicing is the removal of non-coding introns from pre-mRNA and the joining of exons together to create the mRNA that cells use to make proteins. 13
The PRPF4B gene plays an important role in the spliceosome complex, which is a group of molecules that removes noncoding introns of RNA. 14 PRPF4B acts as a serine/threonine kinase within the spliceosome. It adds phosphate groups to specific splicing proteins to regulate their activity. This is required for proper spliceosome assembly, timing and disassembly during the pre-mRNA processing. By doing this, PRPF4B helps to ensure that the introns are correctly removed and the exons are correctly joined. When the PRPF4B kinase activity is changed, it can cause discrepancies in splice-site selections and mRNA creation transcripts. Even though these transcripts could still be expressed at normal levels, it could also encode proteins with impaired function.
Incorrect splicing due to variations within PRPF4B can cause improperly expressed proteins that could disrupt cellular processes that play a role in fluid regulation.15 In the brain, splicing inconsistencies can lead to inaccurate expression of proteins needed for the regulation of fluid transport, ion channels, and cilia function within the ventricular system. 16 This leads to an abnormal accumulation of cerebrospinal fluid and, as a result, variability within intracranial pressures.17 In addition to the brain, PRPF4B splicing errors can impact other organ systems dependent on fluid regulation and correct protein synthesis.18 While these are significantly less minor than CSF regulation in the brain, PRPF4B mutations in other parts of the body can still impact human health. The kidneys, which are responsible for the balance of fluids within the body, may be affected if the incorrect proteins are produced.19, 20 Protein misregulation in the kidneys can affect the functioning of the endothelial cells, which can cause inflammation, oxidative stress, and impaired blood flow in the kidneys.21, 22
Not all genes will respond the same way to the loss of a single allele.23 Some genes are dosage-sensitive. This means that a single functional copy is not enough to maintain normal mRNA and protein levels.24 Others are dosage-tolerant and can compensate through the remaining allele.25 Whether PRPF4B is dosage-sensitive or dosage-tolerant is currently undetermined. Understanding if cells can maintain normal PRPF4B expression with only one functional allele is therefore required for understanding how heterozygous PRPF4B mutations contribute to diseases.
Because patients with PRPF4B-linked neurodevelopmental disorders often have a dominant heterozygous mutation, understanding how reduced PRPF4B function affects fluid production is necessary.26 iPSCs, which are adult cells that have been reprogrammed into a stem-cell-like state, allow researchers to model neural development without needing actual brain tissue from patients.27 It will help researchers understand if a single copy of a functional PRPF4B gene can maintain normal mRNA splicing and protein levels.
Although gene expression is often measured using mRNA levels, lots of mRNA does not always directly correlate to protein levels.28 Post-transcriptional regulation, translation efficiency and protein stability can all impact how much functional protein is synthesized.29 For the PRPF4B gene, mutations could change RNA splicing without changing transcript abundance. Removing the uncertainty through measuring both PRPF4B mRNA levels and PRPF4B protein levels is required to determine whether the loss of an allele affects transcription, protein production or both. This is important for determining whether cells can maintain normal PRPF4B protein levels even with reduced gene expression, which is important for evaluating and understanding the effectiveness of therapeutic strategies that selectively block mutant PRPF4B mRNA.
To understand how disruptions in the PRPF4B gene affect human cells and mRNA splicing, gene editing provides a direct approach.30, 31 CRISPR-Cas9, which stands for Clustered Regularly Interspaced Short Palindromic Repeats Cas9, allows researchers to create targeted gene knockouts.32 This includes creating heterozygous knockouts of PRPF4B to study how reduced protein levels impact cell function and cerebrospinal fluid.
The current study therefore focuses on evaluating PRPF4B protein levels in iPSCs after CRISPR-Cas9–mediated gene removal and measuring its impact on mRNA and protein production. Understanding whether cells can tolerate the loss of a single PRPF4B allele without losing the required protein functions is important to understanding the impact of PRPF4B in brain development and kidney function. This research also provides a basis for future work towards a treatment that reduces the impact of PRPF4B gene mutations in the development of neurodevelopmental disorders.
Having this background, the primary research question of this study is whether human induced pluripotent stem cells (iPSCs) can maintain the normal levels of PRPF4B protein and mRNA expression (normal levels found in wild-type cells) after one allele of the PRPF4B gene is knocked out using CRISPR-Cas9. It is hypothesized that the knockout of a single PRPF4B allele will not affect the level of PRPF4B protein or PRPF4B mRNA compared with non-edited, wild-type PRPF4B cells. This is because the remaining wild-type PRPF4B allele is expected to compensate for the loss by maintaining transcription levels and translation to maintain normal PRPF4B mRNA and protein levels. If this were to not happen within the human body, important organs could be impacted, which could cause development disorders and fluid maintenance problems.
References 1. Wichmann TO, Damkier HH, Pedersen M. A brief overview of the cerebrospinal fluid system and its implications for brain and spinal cord diseases. Front Hum Neurosci. 2021;15:737217. doi:10.3389/fnhum.2021.737217 2. Huff T, Tadi P, Weisbrod LJ, Varacallo M. Neuroanatomy, cerebrospinal fluid. Published online 2017. https://www.ncbi.nlm.nih.gov/books/NBK470578/ 3. Nutter CA, Kidd BM, Carter HA, et al. Choroid plexus mis-splicing and altered cerebrospinal fluid composition in myotonic dystrophy type 1. Brain. 2023;146(10):4217-4232. doi:10.1093/brain/awad148 4. Runge K, Tebartz van Elst L, Maier S, et al. Cerebrospinal fluid findings of 36 adult patients with autism spectrum disorder. Brain Sci. 2020;10(6):355. doi:10.3390/brainsci10060355 5. Jeong J, Yoo HJ, An JY, Jeong S. Dysregulated RNA-binding proteins and alternative splicing: Emerging roles in autism spectrum disorder. Mol Cells. 2025;48(8):100237. doi:10.1016/j.mocell.2025.100237 6. Wang D, Farhana A. Biochemistry, RNA structure. Published online 2020. https://www.ncbi.nlm.nih.gov/books/NBK558999/ 7. Wang Y, Liu J, Huang BO, et al. Mechanism of alternative splicing and its regulation. Biomed Rep. 2015;3(2):152-158. doi:10.3892/br.2014.407 8. Zhou Y, Chen C, Johansson MJO. The pre-mRNA retention and splicing complex controls tRNA maturation by promoting TAN1 expression. Nucleic Acids Res. 2013;41(11):5669-5678. doi:10.1093/nar/gkt269 9. Coelho MC, Pinto RM, Murray AW. Heterozygous mutations cause genetic instability in a yeast model of cancer evolution. Nature. 2019;566(7743):275-278. doi:10.1038/s41586-019-0887-y 10. Simon MJ, Iliff JJ. Regulation of cerebrospinal fluid (CSF) flow in neurodegenerative, neurovascular and neuroinflammatory disease. Biochim Biophys Acta. 2016;1862(3):442-451. doi:10.1016/j.bbadis.2015.10.014 11. Chen X, Liu Y, Sheng X, et al. PRPF4 mutations cause autosomal dominant retinitis pigmentosa. Hum Mol Genet. 2014;23(11):2926-2939. doi:10.1093/hmg/ddu005 12. Tanackovic G, Ransijn A, Thibault P, et al. PRPF mutations are associated with generalized defects in spliceosome formation and pre-mRNA splicing in patients with retinitis pigmentosa. Hum Mol Genet. 2011;20(11):2116-2130. doi:10.1093/hmg/ddr094 13. Will CL, Lührmann R. Spliceosome structure and function. Cold Spring Harb Perspect Biol. 2011;3(7):a003707-a003707. doi:10.1101/cshperspect.a003707 14. Cvitkovic I, Jurica MS. Spliceosome database: a tool for tracking components of the spliceosome. Nucleic Acids Res. 2013;41(Database issue):D132-D141. doi:10.1093/nar/gks999 15. Mathavarajah S, Chipurupalli S, Habib EB, et al. The evolutionarily conserved PRP4K-CHMP4B/vps32 splicing circuit regulates autophagy. Cell Rep. 2025;44(7):115870. doi:10.1016/j.celrep.2025.115870 16. Louvi A, Grove EA. Cilia in the CNS: the quiet organelle claims center stage. Neuron. 2011;69(6):1046-1060. doi:10.1016/j.neuron.2011.03.002 17. Patabendige A, Vinje V, Stoodley M. Editorial: Cerebrospinal fluid dynamics and intracranial pressure elevation-Novel insights on molecular and physiological mechanisms, and implications for neurological disease. Front Mol Neurosci. 2022;15:1119980. doi:10.3389/fnmol.2022.1119980 18. Irwin KE, Jasin P, Braunstein KE, et al. A fluid biomarker reveals loss of TDP-43 splicing repression in presymptomatic ALS-FTD. Nat Med. 2024;30(2):382-393. doi:10.1038/s41591-023-02788-5 19. Zhou X, Lou X, Zhou Y, et al. Novel biallelic mutations in TMEM126B cause splicing defects and lead to Leigh-like syndrome with severe complex I deficiency. J Hum Genet. 2023;68(4):239-246. doi:10.1038/s10038-022-01102-4 20. Whelan SCM, Mutchler SM, Han A, et al. Kcnma1 alternative splicing in mouse kidney: regulation during development and by dietary K+ intake. Am J Physiol Renal Physiol. 2024;327(1):F49-F60. doi:10.1152/ajprenal.00100.2024 21. Diaz-Ricart M, Torramade-Moix S, Pascual G, et al. Endothelial damage, inflammation and immunity in chronic kidney disease. Toxins (Basel). 2020;12(6):361. doi:10.3390/toxins12060361 22. Gyurászová M, Gurecká R, Bábíčková J, Tóthová Ľ. Oxidative stress in the pathophysiology of kidney disease: Implications for noninvasive monitoring and identification of biomarkers. Oxid Med Cell Longev. 2020;2020:5478708. doi:10.1155/2020/5478708 23. Savova V, Vinogradova S, Pruss D, Gimelbrant AA, Weiss LA. Risk alleles of genes with monoallelic expression are enriched in gain-of-function variants and depleted in loss-of-function variants for neurodevelopmental disorders. Mol Psychiatry. 2017;22(12):1785-1794. doi:10.1038/mp.2017.13 24. Eckersley-Maslin MA, Spector DL. Random monoallelic expression: regulating gene expression one allele at a time. Trends Genet. 2014;30(6):237-244. doi:10.1016/j.tig.2014.03.003 25. Mirinezhad MR, Mirzaei F, Salmaninejad A, et al. Reporting a homozygous case of neurodevelopmental disorder associated with a novel PRPF8 variant. Mol Genet Genomic Med. 2025;13(3):e70084. doi:10.1002/mgg3.70084 26. Vadodaria KC, Jones JR, Linker S, Gage FH. Modeling brain disorders using induced pluripotent stem cells. Cold Spring Harb Perspect Biol. 2020;12(6):a035659. doi:10.1101/cshperspect.a035659 27. Payne SH. The utility of protein and mRNA correlation. Trends Biochem Sci. 2015;40(1):1-3. doi:10.1016/j.tibs.2014.10.010 28. Tsoy S, Liu J. Regulation of protein synthesis at the translational level: Novel findings in cardiovascular biology. Biomolecules. 2025;15(5):692. doi:10.3390/biom15050692 29. Hassan MA, Saeij JPJ. Incorporating alternative splicing and mRNA editing into the genetic analysis of complex traits. Bioessays. 2014;36(11):1032-1040. doi:10.1002/bies.201400079 30. Stadelmann C, Di Francescantonio S, Marg A, Müthel S, Spuler S, Escobar H. mRNA-mediated delivery of gene editing tools to human primary muscle stem cells. Mol Ther Nucleic Acids. 2022;28:47-57. doi:10.1016/j.omtn.2022.02.016 31. Gostimskaya I. CRISPR-Cas9: A history of its discovery and ethical considerations of its use in genome editing. Biochemistry (Mosc). 2022;87(8):777-788. doi:10.1134/S0006297922080090
Variables
The independent variable in this study was PRPF4B gene dosage, which was manipulated with CRISPR-Cas9. I compared wild-type iPSCs with two functional PRPF4B alleles to heterozygous PRPF4B knockout iPSCs with one functional allele. The dependent variables were PRPF4B mRNA and protein levels. mRNA levels were measured using RT-qPCR and quantified through Ct values using the 2⁻ΔΔCt method. Protein levels were measured using western blot analysis. To make sure that any differences were because of changes in PRPF4B gene dosage I had to control several variables. Cell culture conditions which consists of temperature (37°C), CO₂ concentration (5%), O₂ concentration (5%), humidity (\~95%), media composition, and passage number range, were kept consistent between wild-type and heterozygous cell cultures. RNA and protein extraction procedures were done using the same kits and protocols. qPCR settings like primer concentrations, thermal cycling parameters, SYBR Green master mix, and instrument settings were kept consistent. Both the RT-qPCR and western blotting procedures were kept consistent by comparing the results in GAPDH, a household gene that retains the same gene expression no matter what happens to the cell.
Procedure
Human induced pluripotent stem cells (iPSCs) were used as a model to test PRPF4B’s dosage sensitivity after heterozygous gene knockout. Both wild-type and heterozygous PRPF4B knockout lines started as single-celled colonies and were incubated for 2 weeks under the exact same conditions. They were kept in defined, serum-free and feeder-free media, with Essential 8 (Thermo Fisher Scientific, Waltham, Massachusetts), at a temperature of 37°C, 5% CO₂ concentration, 5% O₂ concentration, and 95% saturated humidity.
RT-qPCR: Cells were cultured to between 70–90% confluency, where point culture media was removed and cells were washed with phosphate-buffered saline (PBS) to remove residual proteins and debris. Cells were then lysed using a guanidinium-based lysis buffer provided in a commercial RNA extraction kit (Zymo Research, Irvine, CA). This buffer disrupts cell membranes and inactivates RNases. The total RNA is preserved and released. The resulting lysate was processed following the manufacturer’s protocol to separate RNA. To account for differences in cell colony numbers and RNA differences, gene expression was normalized to GAPDH.
To allow for quantification of gene expression, the resulting RNA was reverse-transcribed into complementary DNA (cDNA) using a commercially available cDNA synthesis kit (Zymo Research). This step uses the enzyme reverse transcriptase to create single-stranded complementary DNA (cDNA) from mRNA templates by complementary base pairing. The cDNA starts as a single strand but is duplicated during quantitative PCR. This step is required as RNA is extremely unstable and cannot be consistently amplified by DNA polymerase during PCR.
The obtained cDNA samples were used as templates for real-time quantitative polymerase chain reaction (qPCR). PRPF4B forward (GACTCTTCAGGCACTTCTATCAC) and reverse primers (ACACCTCTCGTAAGTTCATGC) were then added to selectively amplify the amount of PRPF4B DNA in the samples. Primers targeting the GAPDH housekeeping gene (forward: GTCTCCTCTGACTTCAACAGCG, reverse: ACCACCCTGTTGCTGTAGCCAA) were also added because of its stable expression between different experimental conditions.
During qPCR, DNA was amplified by repeating cycles of denaturation, annealing and extension using a CFX96 Real-Time PCR Detection System (Bio-Rad, Hercules, California). The denaturation step, which heats up the cDNA to 95°C, breaks the hydrogen bonds between complementary base pairs. This step is required in order for primers to access the target DNA sequence. During annealing, the temperature was lowered to 65°C to allow forward and reverse primers to bind to the sequences directly around the PRPF4B or GAPDH target spot. Primers determine which part of the DNA strand gets copied and bonds through hydrogen bonds. This temperature must be maintained as it is the temperature in which bonds between the primers and the DNA strands will form at the perfect location for base pairing. These primers dissociate at the beginning of each cycle during the denaturation step when the hydrogen bonds between each DNA strand is broken. Lastly, in the process of extension (at 72°C), DNA polymerase extended from the bound primers synthesizes new DNA strands by adding complementary nucleotides in the 5’ to 3’ direction. This happens to every single place where the primers can be found and creates double-stranded DNA copies of the target region. PowerTrack SYBR Green Master Mix for qPCR (Thermofisher, Waltham, MA) bonds to any dsDNA (double-stranded DNA) that is created by the end of extension. Because double-stranded DNA has stacked base pairs, a major groove and a minor groove. SYBR Green is correct width for the minor groove and has complementary charge distribution, making it able to slide between the adjacent base pairs. In a process called intercalation, the dye is stabilized through LDFs. This allows for fluorescence to be an accurate a measurement of the amount of DNA strands after each cycle. These three steps were repeated until fluorescence reached a certain threshold. The amount of cycles required was recorded.
RT-qPCR Data Analysis: As PCR progressed, greater amounts of double-stranded DNA gets produced, which allows SYBR green dye molecules to emit greater fluorescence. SYBR bonds to the grooves in double-stranded DNA through bouncing around at immense concentrations. These molecules are positively charged compare to the negatively charged DNA strands and fit perfectly in the grooves. When being read by the PCR machine, they jump to a different energy level and transmit small amounts of light. This is then detected by the machine and recorded after every single cycle.
A fluorescence threshold was set in order to determine cycle threshold (Ct) value for each sample. The Ct value represents the number of amplification cycles for the fluorescence to pass this threshold. A sample with higher amounts of starting PRPF4B mRNA would have a lower Ct value while samples with less starting mRNA have a higher Ct value.
To control for differences in RNA input and reverse transcription efficiency, PRPF4B Ct values were normalized to Ct values of GAPDH, the housekeeping gene. This process of normalization was done through the 𝚫Ct equation with both wild-type and heterozygous: 𝚫Ct = Average Ct (PRPF4B) - Average Ct (GAPDH)
To further compare gene expression between heterozygous PRPF4B knockout cells and wild-type, 𝚫Ct values were normalized again using the 𝚫𝚫Ct method: 𝚫𝚫Ct = 𝚫Ct (PRPF4B Heterozygous) - 𝚫Ct (PRPF4B Wild-Type)
Because PCR amplification doubles each cycle and therefore, is exponential, differences in Ct values mean differences in the starting amount of PRPF4B mRNA. Due to Ct values being the amount of cycles needed to reach a threshold, directly comparing Ct values cannot be done. The following equation turns Ct differences into relative expression: 2-𝚫𝚫Ct
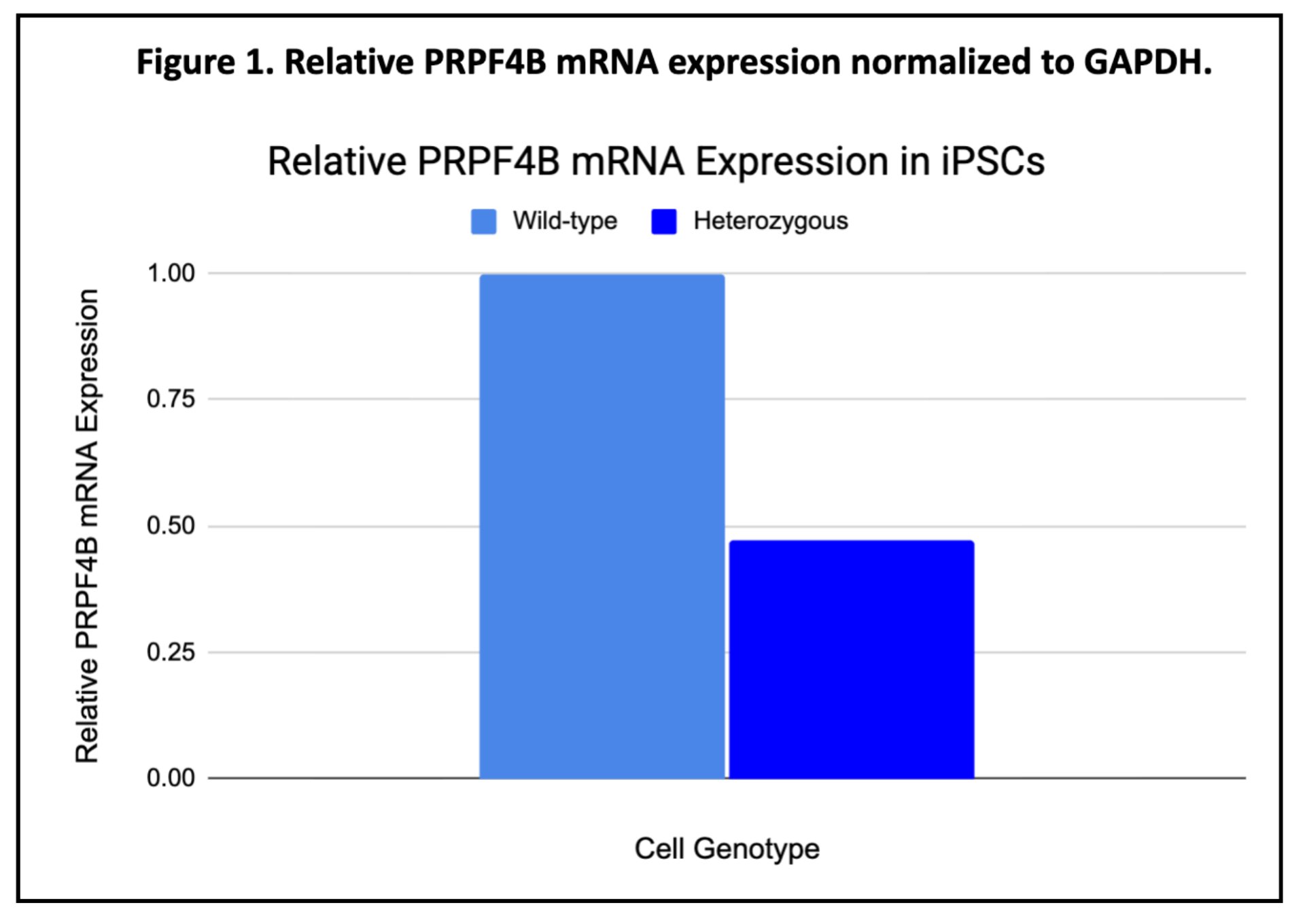
The values represent the relative PRPF4B mRNA expression of heterozygous knockout cells compared to wild-type cells, having wild-type expression as the reference value. A value of 1 means there was no change found in expression relative to wild-type, values less than 1 show less PRPF4B expression in heterozygous cells and values greater than 1 mean increased PRPF4B expression relative to wild-type. Because wild-type samples are used as the control, they represent the maximum normalized expression level, and all heterozygous values are a proportion of wild-type expression.
Western Blotting: To determine whether PRPF4B protein levels remain constant after heterozygous gene knockout, quantitative Western blotting was used on wild-type and heterozygous PRPF4B knockout human induced pluripotent stem cell (iPSC) populations. After the identical processes of incubation, the cells were harvested and lysed using a protein extraction buffer with detergents and protease inhibitors. This step destroyed cellular membranes and released intracellular proteins, avoiding enzymatic degradation in the process.
After lysis, protein lysates were centrifuged to remove cellular debris. Equal amounts of total protein lysate from each sample was then prepared for electrophoresis. Any variation in protein loading was controlled during data analysis by normalizing PRPF4B protein expression to GAPDH protein expression. Protein samples were mixed with sodium dodecyl sulfate (SDS) sample buffer and heated to denature protein secondary and tertiary structures. SDS binds to proteins and gives them a consistently negative charge, which allows proteins to migrate toward the positive electrode during electrophoresis in the following step. Denaturation makes sure all proteins are separated only on molecular weight.
Denatured protein samples were separated in a sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). A stacking gel and resolving gel were employed to allow for consistent protein separation. The stacking gel concentrated proteins into narrow bands, which allowed for uniform protein migration. The resolving gel separated proteins based on molecular weight.
After electrophoresis, separated proteins where transferred from the gel onto a polyvinylidene difluoride (PVDF) membrane through electrotransfer. This process stopped the movement of proteins on the membrane and allowed for antibody-based quantification of these proteins. The membrane was then incubated into a blocking solution containing non-fat milk proteins to prevent non-specific antibody binding.
A Novus Biologicals NB100-86997 primary antibody was then incubated with the membrane to bind with PRPF4B proteins while a Sigma G9545 rabbit antibody that targeted GAPDH was used to detect GAPDH proteins as a housekeeping protein. These antibodies selectively bind to their target protein because of antigen-antibody specificity.
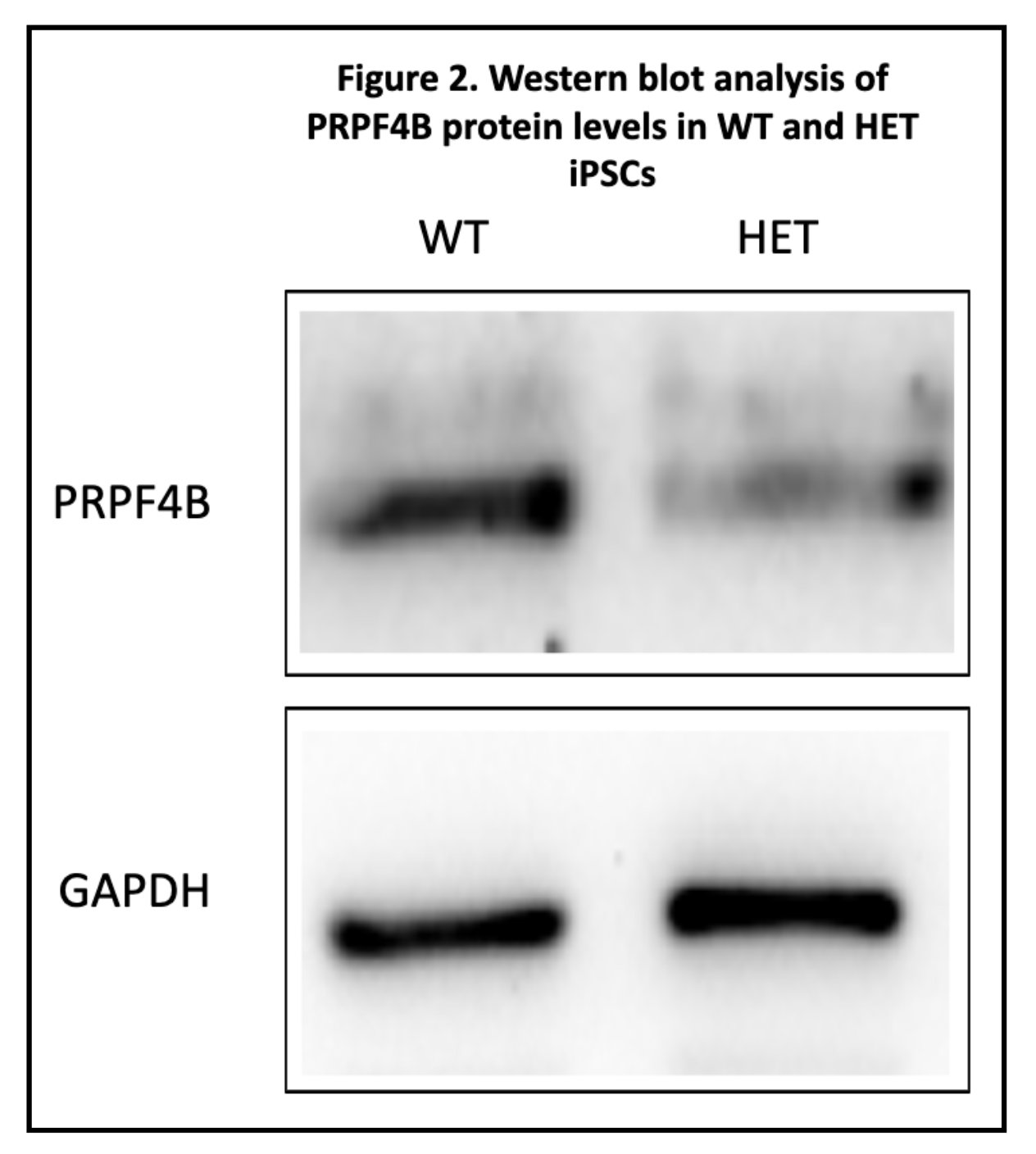
After washing to remove unbound primary antibodies, the membranes were incubated with a secondary antibody. The second antibody was conjugated to a chemiluminescent enzyme reporter. When exposed to chemiluminescent substrate, the enzyme creates a light-producing reaction at antibody binding sites. The emitted light signal was seen and measured using an imaging system. Because light intensity corresponds to protein amount, the band intensities could be directly compared.
Observations
Through a Western Blot as well as RT-qPCR, I saw less mRNA and less proteins. Both observations can be seen through the graphs/tables below.
Analysis
Heterozygous PRPF4B knockout (with only one single functional allele) had less PRPF4B mRNA and protein levels compared to wild-type cells. This shows that a single functional allele is insufficient to maintain normal expression levels, meaning that PRPF4B is dosage-sensitive. The remaining functional allele cannot compensate through increased expression. Understanding that something is going wrong during the transcription stage, this allows for more targeted solutions in the future.
Conclusion
PRPF4B-linked disorders may be because of insufficient gene expression. This means that knocking out an allele to create heterozygous KO would lead to less mRNA and less proteins. A potential strategy could be restoring PRPF4B expression by enhancing transcription from the remaining allele through activating the PRPF4B promoter or using transcription-enhancing drugs.
Application
Reduced PRPF4B expression is a contributor for neurodevelopment disorders in certain cases as it messes with normal RNA splicing and gene regulation. Understanding PRPF4B dosage sensitivity can help identify mechanisms underlying these conditions and can help create therapies that restore normal gene expression like potentially applying transcription enhancing drugs as well as a PRPF4B promoter.
Sources Of Error
There were two major sources of error in this project. One being the use of iPSCs. iPSCs, or human induced pluripotent stem cells rather than actual brain cells. Although iPSCs have been found in past studies to well represent the brain, it doesn't act the exact same. Due to major time constraints as well as difficulties accessing brain cells, they were not used. Additionally, protein levels were analyzed rather than protein functionality. Protein functionality measures how usable the proteins are to the cell while protein levels only measures how much, meaning that the protein level also includes non-functional protesin manufactured. Fixing this step to include functionality would be a strong advancement for this project.
Citations
References 1. Wichmann TO, Damkier HH, Pedersen M. A brief overview of the cerebrospinal fluid system and its implications for brain and spinal cord diseases. Front Hum Neurosci. 2021;15:737217. doi:10.3389/fnhum.2021.737217 2. Huff T, Tadi P, Weisbrod LJ, Varacallo M. Neuroanatomy, cerebrospinal fluid. Published online 2017. https://www.ncbi.nlm.nih.gov/books/NBK470578/ 3. Nutter CA, Kidd BM, Carter HA, et al. Choroid plexus mis-splicing and altered cerebrospinal fluid composition in myotonic dystrophy type 1. Brain. 2023;146(10):4217-4232. doi:10.1093/brain/awad148 4. Runge K, Tebartz van Elst L, Maier S, et al. Cerebrospinal fluid findings of 36 adult patients with autism spectrum disorder. Brain Sci. 2020;10(6):355. doi:10.3390/brainsci10060355 5. Jeong J, Yoo HJ, An JY, Jeong S. Dysregulated RNA-binding proteins and alternative splicing: Emerging roles in autism spectrum disorder. Mol Cells. 2025;48(8):100237. doi:10.1016/j.mocell.2025.100237 6. Wang D, Farhana A. Biochemistry, RNA structure. Published online 2020. https://www.ncbi.nlm.nih.gov/books/NBK558999/ 7. Wang Y, Liu J, Huang BO, et al. Mechanism of alternative splicing and its regulation. Biomed Rep. 2015;3(2):152-158. doi:10.3892/br.2014.407 8. Zhou Y, Chen C, Johansson MJO. The pre-mRNA retention and splicing complex controls tRNA maturation by promoting TAN1 expression. Nucleic Acids Res. 2013;41(11):5669-5678. doi:10.1093/nar/gkt269 9. Coelho MC, Pinto RM, Murray AW. Heterozygous mutations cause genetic instability in a yeast model of cancer evolution. Nature. 2019;566(7743):275-278. doi:10.1038/s41586-019-0887-y 10. Simon MJ, Iliff JJ. Regulation of cerebrospinal fluid (CSF) flow in neurodegenerative, neurovascular and neuroinflammatory disease. Biochim Biophys Acta. 2016;1862(3):442-451. doi:10.1016/j.bbadis.2015.10.014 11. Chen X, Liu Y, Sheng X, et al. PRPF4 mutations cause autosomal dominant retinitis pigmentosa. Hum Mol Genet. 2014;23(11):2926-2939. doi:10.1093/hmg/ddu005 12. Tanackovic G, Ransijn A, Thibault P, et al. PRPF mutations are associated with generalized defects in spliceosome formation and pre-mRNA splicing in patients with retinitis pigmentosa. Hum Mol Genet. 2011;20(11):2116-2130. doi:10.1093/hmg/ddr094 13. Will CL, Lührmann R. Spliceosome structure and function. Cold Spring Harb Perspect Biol. 2011;3(7):a003707-a003707. doi:10.1101/cshperspect.a003707 14. Cvitkovic I, Jurica MS. Spliceosome database: a tool for tracking components of the spliceosome. Nucleic Acids Res. 2013;41(Database issue):D132-D141. doi:10.1093/nar/gks999 15. Mathavarajah S, Chipurupalli S, Habib EB, et al. The evolutionarily conserved PRP4K-CHMP4B/vps32 splicing circuit regulates autophagy. Cell Rep. 2025;44(7):115870. doi:10.1016/j.celrep.2025.115870 16. Louvi A, Grove EA. Cilia in the CNS: the quiet organelle claims center stage. Neuron. 2011;69(6):1046-1060. doi:10.1016/j.neuron.2011.03.002 17. Patabendige A, Vinje V, Stoodley M. Editorial: Cerebrospinal fluid dynamics and intracranial pressure elevation-Novel insights on molecular and physiological mechanisms, and implications for neurological disease. Front Mol Neurosci. 2022;15:1119980. doi:10.3389/fnmol.2022.1119980 18. Irwin KE, Jasin P, Braunstein KE, et al. A fluid biomarker reveals loss of TDP-43 splicing repression in presymptomatic ALS-FTD. Nat Med. 2024;30(2):382-393. doi:10.1038/s41591-023-02788-5 19. Zhou X, Lou X, Zhou Y, et al. Novel biallelic mutations in TMEM126B cause splicing defects and lead to Leigh-like syndrome with severe complex I deficiency. J Hum Genet. 2023;68(4):239-246. doi:10.1038/s10038-022-01102-4 20. Whelan SCM, Mutchler SM, Han A, et al. Kcnma1 alternative splicing in mouse kidney: regulation during development and by dietary K+ intake. Am J Physiol Renal Physiol. 2024;327(1):F49-F60. doi:10.1152/ajprenal.00100.2024 21. Diaz-Ricart M, Torramade-Moix S, Pascual G, et al. Endothelial damage, inflammation and immunity in chronic kidney disease. Toxins (Basel). 2020;12(6):361. doi:10.3390/toxins12060361 22. Gyurászová M, Gurecká R, Bábíčková J, Tóthová Ľ. Oxidative stress in the pathophysiology of kidney disease: Implications for noninvasive monitoring and identification of biomarkers. Oxid Med Cell Longev. 2020;2020:5478708. doi:10.1155/2020/5478708 23. Savova V, Vinogradova S, Pruss D, Gimelbrant AA, Weiss LA. Risk alleles of genes with monoallelic expression are enriched in gain-of-function variants and depleted in loss-of-function variants for neurodevelopmental disorders. Mol Psychiatry. 2017;22(12):1785-1794. doi:10.1038/mp.2017.13 24. Eckersley-Maslin MA, Spector DL. Random monoallelic expression: regulating gene expression one allele at a time. Trends Genet. 2014;30(6):237-244. doi:10.1016/j.tig.2014.03.003 25. Mirinezhad MR, Mirzaei F, Salmaninejad A, et al. Reporting a homozygous case of neurodevelopmental disorder associated with a novel PRPF8 variant. Mol Genet Genomic Med. 2025;13(3):e70084. doi:10.1002/mgg3.70084 26. Vadodaria KC, Jones JR, Linker S, Gage FH. Modeling brain disorders using induced pluripotent stem cells. Cold Spring Harb Perspect Biol. 2020;12(6):a035659. doi:10.1101/cshperspect.a035659 27. Payne SH. The utility of protein and mRNA correlation. Trends Biochem Sci. 2015;40(1):1-3. doi:10.1016/j.tibs.2014.10.010 28. Tsoy S, Liu J. Regulation of protein synthesis at the translational level: Novel findings in cardiovascular biology. Biomolecules. 2025;15(5):692. doi:10.3390/biom15050692 29. Hassan MA, Saeij JPJ. Incorporating alternative splicing and mRNA editing into the genetic analysis of complex traits. Bioessays. 2014;36(11):1032-1040. doi:10.1002/bies.201400079 30. Stadelmann C, Di Francescantonio S, Marg A, Müthel S, Spuler S, Escobar H. mRNA-mediated delivery of gene editing tools to human primary muscle stem cells. Mol Ther Nucleic Acids. 2022;28:47-57. doi:10.1016/j.omtn.2022.02.016 31. Gostimskaya I. CRISPR-Cas9: A history of its discovery and ethical considerations of its use in genome editing. Biochemistry (Mosc). 2022;87(8):777-788. doi:10.1134/S0006297922080090
Acknowledgement
Throughout this project, I had several mentors aiding me through primarily the understanding portion. While all the experiments and writting have been done by myself, I would still like to thank Dr. Heng, Dr. Yang as well as Dr. Garcia for their help throughout this project.