
Dancing Molecules: Adaptive Scaffolds for Tissue Regeneration
Bhagat Hithesh, Sarbajit Sen
Tom Baines School
Grade 8
Presentation
Problem
1.1: Tissue Healing Overview
When the human body heals from an injury, its tissues don’t just build up and try to close the wound, they have to reorganize, realign, and respond depending upon movement/mechanical forces. This is an extremely crucial part of rebuilding and replenishing tissues, bones, nerves and blood vessels. The initial phase of healing, known as the ‘Granulation/Proliferative Phase’ begins 3 days after an injury. When new tissue starts to form, the wound contracts and new blood vessels emerge. During this phase, extracellular environment and the mechanical forces acting on cells play a critical role in determining whether tissue growth is organized or disordered.
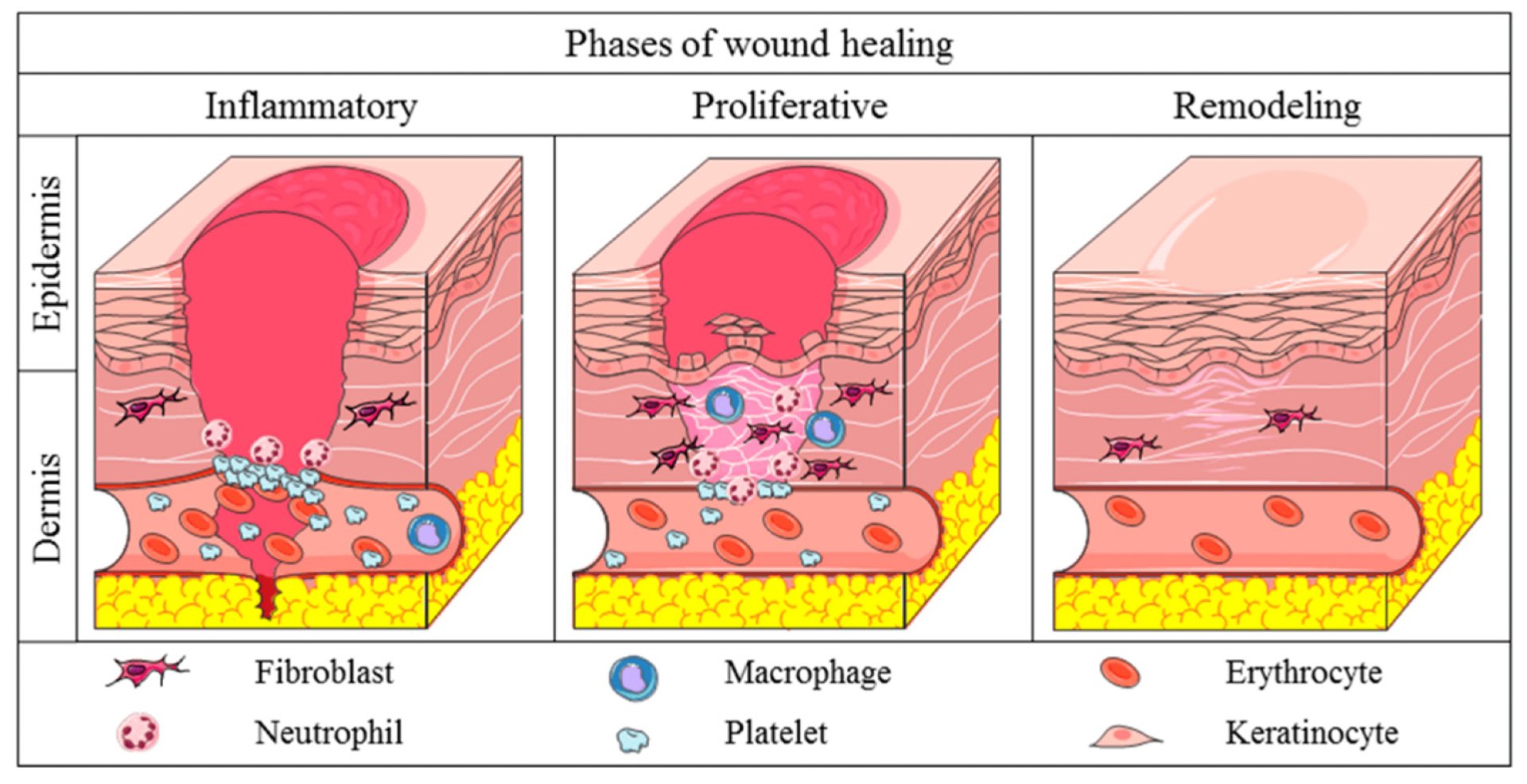 Image courtesy: Alberts, A., Lungescu, I. A., Niculescu, A.-G., & Grumezescu, A. M. (2025). Natural Products for Improving Soft Tissue Healing: Mechanisms, Innovations, and Clinical Potential. Pharmaceutics, 17(6), 758
Image courtesy: Alberts, A., Lungescu, I. A., Niculescu, A.-G., & Grumezescu, A. M. (2025). Natural Products for Improving Soft Tissue Healing: Mechanisms, Innovations, and Clinical Potential. Pharmaceutics, 17(6), 758
1.1.1: Axonal Damage
However when the damage forms, nerve fibers known as axons are severed, disrupting communication to the brain as axons are responsible for transmitting electrical signals along the central nervous system. When severed, these signals cannot travel effectively, resulting in loss of sensation, impaired motor function, or complete paralysis, depending upon the location and level of the injury.
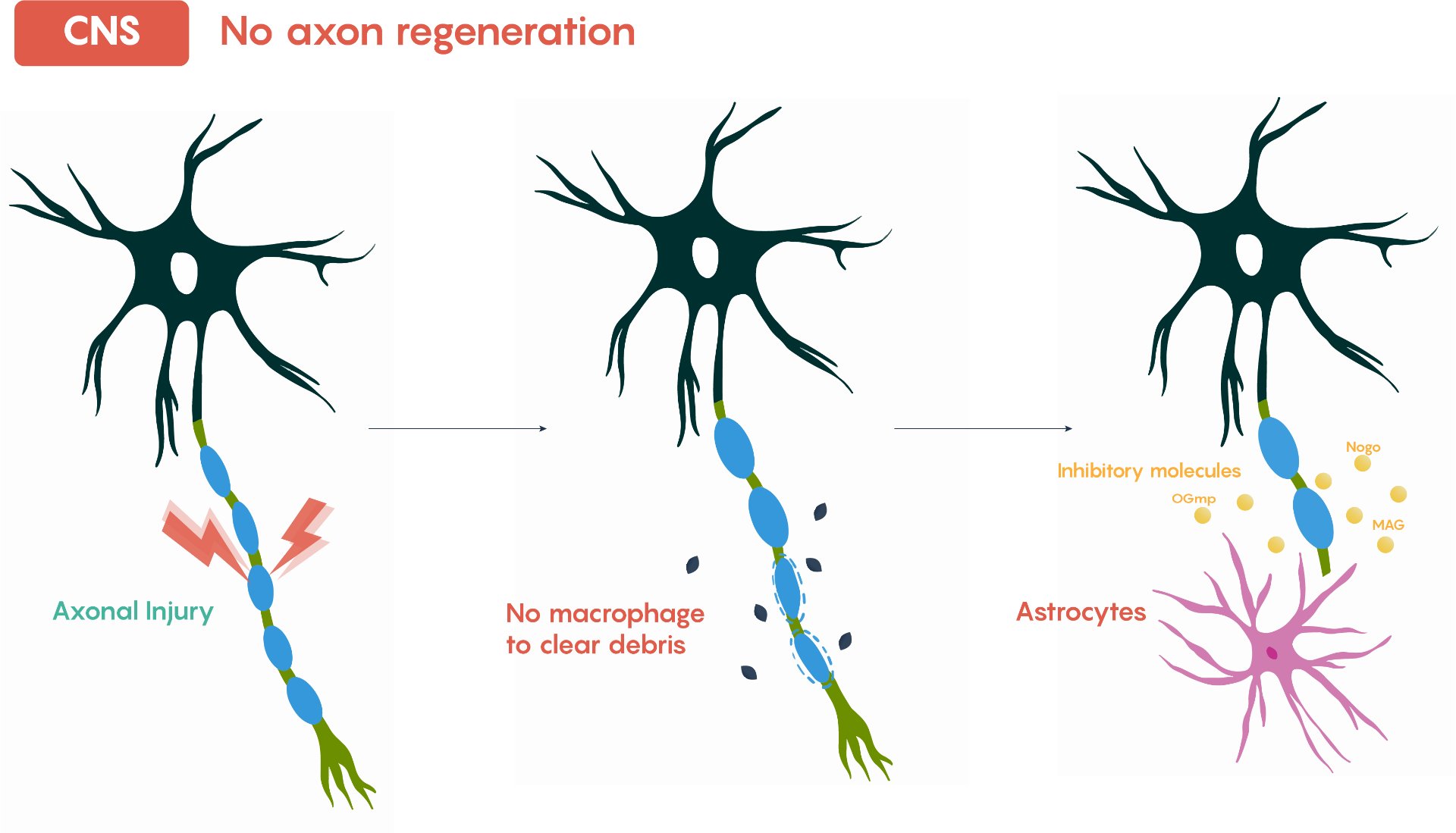 Image Courtesy: Ju, W., Santos, A., Freeman, A., & Daniele, E. (2018). Neuroscience: Canadian 1st Edition [Open textbook]. eCampusOntario.
Image Courtesy: Ju, W., Santos, A., Freeman, A., & Daniele, E. (2018). Neuroscience: Canadian 1st Edition [Open textbook]. eCampusOntario.
1.1.2: Inflammation and Glial Scarring
The body responds to spinal injuries with inflammation to prevent infection and initiate repair. In this process, astrocytes proliferate and secrete components of the extracellular matrix, creating a dense physical barrier called a glial scar. While the scar prevents further tissue damage, it blocks axonal regrowth, preventing neurons from reconnecting across the injury site. A major consequence is the loss of myelin, the insulating layer around neurons that ensures efficient electrical signal transmission, Without myelin, even surviving neurons are less effective at conducting signals, contributing to paralysis.
1.1.3: Types of Paralysis
Types of paralysis caused by spinal injuries include...
| Type of Paralysis | Description |
|---|---|
| Paraplegia | When the spinal cord is damaged in the thoracic or lumbar region, the individual is not able to move or feel sensations in the lower limbs, making it extremely difficult or impossible for them to walk, stand, or move the legs. Besides the motor effects, the individual may also experience some loss of bladder, bowel, blood pressure, and temperature control, as the spinal cord is also damaged, thus affecting the autonomic nervous system (Chen et al., 2022; Silver & Miller, 2004; Slotkin et al., 2017). |
| Quadriplegia (Tetraplegia) | Damage to the cervical part of the spinal cord leads to partial or total loss of movement and sensation in all four limbs, including the arms, hands, trunk, and legs. At this level, there is also impairment in the ability to breathe, swallow, control the head and neck, and the bladder and bowel, among others. The extent to which this occurs depends on the level and completeness of the injury. Recovery can also not occur without the use of advanced medical techniques such as regenerative scaffold techniques, nerve guidance therapies, and rebuild techniques. |
| Partial/Incomplete | However, not all spinal cord injuries result in complete paralysis. In incomplete spinal cord injuries, some nerve signals still get through the injury site. These signals enable partial function below the injury site. Example: If a person is an incomplete paraplegic, it means they still have some function in one of their legs. If they are an incomplete quadriplegic, it means they still have some function in their hands. The extent of this function depends on the extent of axonal preservation, level of injury, and success of nerve regeneration strategies. |
1.2: Molecular Dynamics in Tissue Healing
In natural biological systems, tissue healing and regeneration occur across multiple scales, from macroscopic tissue remodeling down to molecular motion. Proteins, extracellular matrix components, and signaling molecules are never static - they exhibit vibrational, rotational, and translational motions driven by thermal energy and Brownian dynamics. This continuous movement leads to frequent molecular collisions, which are essential for interactions such as receptor binding, mechanosignaling, and structural organization (Frantz, Stewart, & Weaver 2010; Magani et al., 2025). According to collision theory, the effectiveness of interaction increases when molecules can move freely, reorient, and adapt - a principle foundational to biological function and molecular communication.
1.2.1: Motion-Driven Adaptability
Living tissue rely on this dynamic behaviour. Structural proteins like collagen, laminin, and fibronectin are not rigid; they dynamically reconfigure in response to mechanical forces, enabling cells to sense strain, adjust orientation, and promote organized growth after injury (Mangani et al., 2025). This motion-driven adaptability enables tissue to regain structure, strength and alignment following trauma - for example, guiding axons during nerve regeneration or remodeling the extracellular matrix during wound healing.
1.2.2: limitations of Static Scaffolds
However, in many synthetic tissue engineering systems, scaffolds are designed with fixed architecture and rigid internal structures, prioritizing static strength over dynamic behaviour. These rigid designs restrict molecular freedom, limiting the ability of scaffold components to reorient or self-correct in response to mechanical stresses. As a consequence, interaction efficiency can be reduced, mechanotransduction signaling is constrained, and adaptive realignment that is critical in living tissue healing tissue is lost. When static scaffolds are stretched, deformed, or misaligned, they are unable to reorganizes their internal structure in ways that sustain prolonged regenerative processes or support recovery of complex injuries. This limitation has profound implications for spinal cord injuries (SCI), which are a major clinical problem worldwide. The spinal cord, part of the central nervous system, is divided into regions with distinct functions.
1.2.3: Spinal Cord Anatomy
Distinctive Regions (Spine) Notes Cervical Spine (C1-C8) Controls neck, arm, and hand function; injuries often cause Quadriplegia. Thoracic Spine (T1-T12) Controls partial leg movement; injuries typically result in paraplegia. Lumbar Spine (L1-L5) Supports lower limb function and sensation; injuries may cause lower body paralysis. Sacral Spine (S1-S5) Involved in bladder, bowel, and sexual function; injuries affect these autonomic processes.
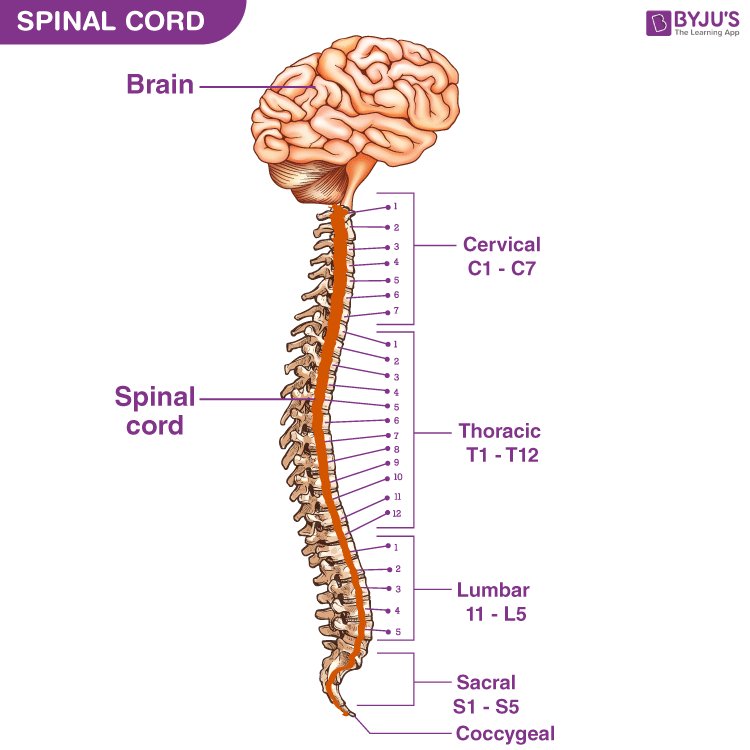
Image Courtesy: https://byjus.com/biology/spinal-cord/
This results in a complex interplay that includes inflammation, cell death, and glial scar tissue, which limits the regeneration of the damaged areas in the spinal cord (Chen et al., 2022; Neural Regeneration Research, 2025). However, scaffolds can provide a bridge that can span the damaged area in the spinal cord. However, there are limitations that still need to be overcome, such as the integration of the scaffolds into the damaged area without causing foreign body reactions, the development of vascular tissue, the limitation of inhibitory scarring, and the alignment of the axons along the appropriate spinal tracts (Zheng et al., 2024; Slotkin et al., 2017).
1.3: Past Approaches to Spinal Cord Regeneration
1.3.1: Acute Clinical Interventions
Acute management of spinal cord injury (SCI) primarily focuses on limiting secondary damage and stabilizing the patient rather than directly restoring neural connectivity. Surgical decompression is often performed to relieve mechanical pressure on the spinal cord caused by fractured vertebrae, hemorrhage, or swelling. By reducing compression, surgeons aim to minimize further neuronal death and ischemic injury.
Historically, high-close corticosteroids such as methylprednisolone were administered to reduce inflammation and limit oxidative stress during the early post-injury period. Although controversial in recent years due to side effects and variable efficacy, corticosteroids were widely used to mitigate aspects of secondary injury. Spinal stabilization procedures, including internal fixation with rods or screws, are used to prevent further mechanical disruption and maintain alignment of the vertebral column.
Following acute care, rehabilitation therapy plays a crucial role in maximizing functional recovery through neuroplastic adaptation, muscle strengthening, and compensatory training. While these interventions can reduce secondary injury progression, they do not directly regenerate severed axonal pathway or re-establish lost synaptic connections across the lesion site.
As a result, permanent deficits often persist, highlighting the need for regenerative strategies that actively promote neural connection.
1.3.2: Growth Factor & Molecular Therapies
Beyond acute interventions, experimental strategies have focused on supplementing or restoring biochemical signals that support neural survival and regeneration. Key molecular approaches include the administration of neurotrophic factors. The molecules act by binding to specific receptors on neurons and glial cells, triggering intracellular signaling pathways that enhance survival, guide axonal growth, and modulate local extracellular matrix remodeling. In principle, creating concentration gradients of growth factors can direct axonal regeneration.
Despite their promise, these molecular therapies face several challenges:
Rapid Degradation - many neurotrophic factors are quickly broken down in vivo\, limiting their effective duration.
Diffusion Constraints - Static delivery systems often fail to maintain stable gradients over the injury site, reducing the guidance efficiency for axons.
Limited integration with tissue mechanics - molecular therapies alone do not provide structural support or respond adaptively to mechanical forces in the spinal cord, which are critical for guiding regenerating axons.
Together, these limitations indicate that while molecular supplementation can support neuronal growth, effective regeneration requires strategies that integrate biochemical guidance with structural and dynamic cues - a need addressed in part by motion - responsive scaffold designs. The poster below is a visual created by us and it focuses on Growth Factor & Molecular Therapies in the context of Neural Regeneration.

1.3.3: Cell-Based Therapies
Cell-based approaches aim to replace lost or damaged neurons, provide growth-supportive environments, or modulate inhibitory signals at the injury site. Examples include:
| Cell Type | Cell Description | Challenges & Limitations |
|---|---|---|
| Stem Cells | Can differentiate into neurons, glial cells, or supportive matrix components. Neural Progenitor Cells Pre-differentiated cells intended to integrate into spinal tissue and form functional synaptic connections. |
|
| Neural Progenitor Cells | Pre-differentiated cells intended to integrate into spinal tissue and form functional synaptic connections. | Same as above |
| Schwann cells & Olfactory Ensheathing cells | Support axonal regrowth and remyelination. | Same as above |
1.3.4: Reframing Scaffold Designs
Scaffolds can provide structural support and cell adhesion, but the lack of motion responsiveness in the scaffolds can be a major drawback in the regeneration process. Motion responsiveness is a key factor in the regeneration process, as it can provide the appropriate guidance to the cells along the complex spinal tracts, such as the corticospinal tracts, which are responsible for the voluntary movement of the limbs, and the dorsal columns, which are responsible for the sensory input.
Therefore, the key scientific problem this project addresses is: how can molecular systems can be designed to intentionally utilize molecular motion - "dancing" - to improve interaction efficiency, alignment recovery, and self-restoration in biomimetic scaffolds? If this could be achieved, it would represent a step toward adaptive healing systems that closely emulate the behaviour of living tissues, with the potential to restore function after injuries at specific spinal levels and reduce paralysis outcomes.
To further explore this, our project investigates the concept of "dancing molecules," a framework in which internal motion - including vibrational, rotational, and translational movement - enables continuous reorientation, adaptive interaction, and shape recovery under varying forces.
By comparing motion-enabled dynamics to traditional rigid scaffolds, this research aims to establish a foundational understanding of how motion-driven design can lead to more effective, adaptive, and biomimetic healing systems inspire by nature's own regenerative strategies.
Method
2.1: Background & Conceptual Framework
Regenerative biology recognizes tissue repair as a highly dynamic and mechanically regulated process. Healing is governed not only by biochemical signalling cascades but also by spatial organization, mechanical strain, and structural adaptability within the extracellular environment. In tissue engineering, a scaffold is defined as a three-dimensional structural framework that provides mechanical support and biochemical guidance to facilitate cell adhesion, proliferation, migration, and differentiation.
In native systems, this structural role is fulfilled by the extracellular matrix (ECM), a dynamic network of proteins such as collagen, laminin, and fibronectin that undergoes continuous remodelling in response to cellular traction forces and biochemical stimuli (Frantz et al., 2010). The ECM actively participates in mechanotransduction—the process by which cells convert mechanical stimuli into biochemical signals (Ingber, 2006).
Unlike static artificial scaffolds, native ECM exhibits viscoelastic behaviour, micro-scale fibre rearrangement, and adaptive reorientation under mechanical stress. These dynamic properties influence cytoskeletal alignment, axonal extension, angiogenesis, and cellular polarity.
This project is grounded in the hypothesis that mechanical micro-movement itself acts as a biological signal. By engineering a scaffold capable of oscillation, realignment, and internal micro-dynamic restructuring—conceptualized as the "Dancing Molecules Effect"—it may be possible to more accurately replicate the regenerative microenvironment of living tissue following spinal trauma.
2.2: Theoretical Rationale
2.2.1: Molecular Signaling in Spinal Injury Repair
Following spinal cord injury, a complex cascade of molecular events regulates inflammation, axonal degeneration, neuroprotection, and regeneration (Silver & Miller, 2004).
Key molecular mediators include:
| Type | Example | Function |
|---|---|---|
| Growth Factors | BDNF (Brain-Derived Neurotrophic Factor), NGF (Nerve Growth Factor), and VEGF (Vascular Endothelial Growth Factor). | Stimulate neuron survival, growth, and blood vessel simulation. |
| Cytokines | IL-6, TNF-α, and IL-10 (these cytokines above drive inflammation). | Regulates the amount of inflammation |
| Extracellular Matrix Proteins | Laminin, Fibronectin, and Collagen | Give structural scaffolds and guide axon alignment. |
| Peptides | Short chains of amino acids like RGD motifs. | Help cells attach to materials and trigger regeneration. |
These molecules form concentration gradients that guide cellular migration via chemotaxis. The behaviour of these gradients can be modelled using Fick's Second Law of Diffusion.

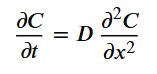 Where:
Where:
- 𝐶C = concentration
- 𝑡t = time
- 𝐷D = diffusion coefficient
In static scaffolds, diffusion is limited by fixed pore geometry, potentially reducing effective 𝐷D. In contrast to this, micro-scale motion may increase local mixing, effectively enhancing diffusion transport.
2.2.2: Static vs Dynamic Scaffold Behaviour
Static Scaffold Limitations:
- Fixed diffusion pathways
- Gradient Stagnation
- Reduced mechanotransductive signaling
- Limited electrical conductivity
- Inability to adapt under mechanical load
Dynamic Scaffold advantages:
- Increase the effective diffusion coefficient through micro-mixing.
- Maintain gradient stability
- Enhance integrin clustering and adhesion
- Improve axonal guidance
- Preserve conductive pathways during deformation
Dynamic mechanical stimulation has been shown to influence neural differentiation and alignment (Discher et al., 2005). Therefore, a scaffold incorporating oscillatory behaviour may provide both biochemical and mechanical guidance.
2.2.3: Electrical Signal Restoration
Neural communication relies on action potential propagation along axons. After spinal injury, conduction pathways are interrupted, impairing synaptic connectivity.
Conductive scaffolds incorporating carbon nanotubes or conductive polymers (e.g., polypyrrole) have demonstrated improved neurite outgrowth and electrical coupling.
Electrical conductivity follows Ohm's Law:
I=V/R or V=IR
Where reducing resistance within a scaffold improves signal propagation.
A dynamic scaffold may:
- Increase neuron-material contact surface area.
- Maintain conductive alignment during deformation.
- Reduce microstructural discontinuities.
Thus, dynamic motion may reduce signal attenuation and enhance functional network reconstruction.
2.3: Scientific Motivation & Prior Research
Natural tissue healing involves structural realignment under mechanical loading. Collagen fibres reorganize along stress vectors, restoring tensile integrity (Fratzi, 2008). Mechanical strain influences cytoskeletal tension, gene expression, and ECM remodelling (Ingber, 2006).
Biomimetic scaffolds have been developed to replicate ECM properties (Hollister, 2005). However, many remain static once fabricated. Recent advances include:
- Shape memory polymers
- Self-healing hydrogels
- Stimuli-responsive materials
- Magnetically actuated biomaterials
These materials demonstrate reorientation and structural memory. The proposed "Dancing Molecules Effect" builds upon this literature by suggesting that continuous micro-scale oscillation increases interaction probability and structural resilience.
Understanding how internal dynamic motion affects scaffold alignment may help bridge the gap between synthetic materials and living tissue behaviour.
2.4: Alternative Modelling Method
A transparent microchannel system will simulate injury-like spinal pathways. Two scaffold conditions will be tested:
- Static Scaffold
- Dynamic Oscillatory Scaffold
Measured Variables:
- Diffusion rate (modeled concentration gradients)
- Interaction frequency with receptor zones
- Realignment efficiency post-deformation
- Electrical continuity (simulated conductive network mapping)
Statistical analysis will compare mean interaction frequencies across trials.
2.5: Hypothesis
If a scaffold incorporates internal dynamic motion that allows continuous reorientation and oscillation, then it will demonstrate increased interaction frequency, improved alignment recovery, and enhanced probabilistic binding efficiency compared to a static scaffold analogue. This is because increased mobility raises collision probability and adaptive restructuring under mechanical disturbance.
2.6: Purpose
The purpose of this project is to investigate whether the dynamic internal movement (model of a “Dancing Molecule” effect) can improve the alignment and self-restoring ability of the biomimetic scaffold model. By comparing the dynamic model to the static one, it would be easier to explore how the recovery after a force is influenced. Thus, we would be able to tackle a broader question: Can movement-responsive scaffolds better mimic natural tissue healing behaviour?
2.7: Conceptual Model/Simulation
To efficiently show the comparisons between rigid molecules and dancing molecules, we have designed a computer-based model that visualizes the movement.
 The molecular agents are separated with the spinal receptors in the middle.
Dancing molecules in red show distinctive vibrations and move at a faster pace; ergo, the connections made are faster than those of traditional rigid molecular agents.
We count the connections based on the duration of time. In five trials with the same time duration, we can understand in a deeper way how molecular motion works in this scenario. Even though the results would be random each time we run it, we would find an average in our data. To further enhance our modelling, we will create a physical model of this experiment and then repeat the same process to compare both physical and computational simulations.
The molecular agents are separated with the spinal receptors in the middle.
Dancing molecules in red show distinctive vibrations and move at a faster pace; ergo, the connections made are faster than those of traditional rigid molecular agents.
We count the connections based on the duration of time. In five trials with the same time duration, we can understand in a deeper way how molecular motion works in this scenario. Even though the results would be random each time we run it, we would find an average in our data. To further enhance our modelling, we will create a physical model of this experiment and then repeat the same process to compare both physical and computational simulations.
(We created this simulation with the help of some ai chatbots, Chatgpt, Gemini, etc.)
2.8 Physical Models
2.8.1: Pendulum-based Model
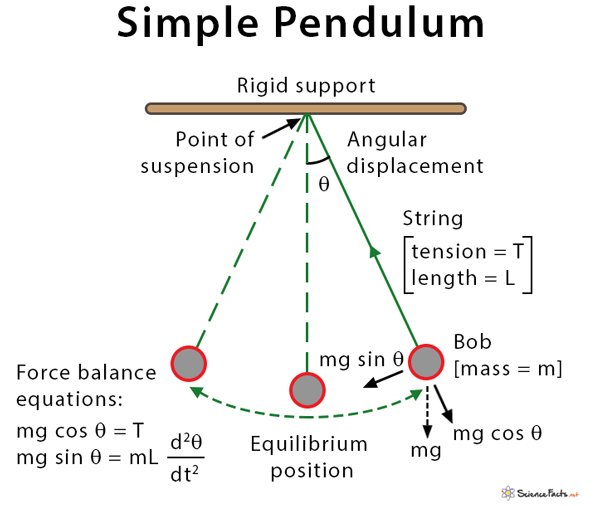
In order to visually demonstrate that “dancing molecule” effect, our project will focus on a physical, pendulum-based model rather than primarily relying on the computational model—one that would support illustration of key concepts of molecular motion and adaptive interaction.
The structure of this physical model would consist of two regions that are separated by a central “spinal receptor” zone. The molecular agents would be rigid and dancing molecules. Rigid ones are going to be represented by stationary pendulums, and that demonstrates motion but slightly limited.
On the other hand, dancing molecules would be represented by double-joined pendulums that oscillate, generally showing a higher amount of kinetic energy as well as continuous reorientation.
Demonstration mechanics of the model are lateral and vibrational movements of the pendulums that would allow the dancing molecules to interact with central receptors more frequently, while rigid ones would get fewer interactions. Connections can be visually represented by magnets, like when a pendulum reaches the receptor zone. Multiple trials would allow for observation on patterns, interaction, and alignment, which judges our physical model on its overall accuracy.
 This is a picture taken of our moving double pendulum in motion.
This is a picture taken of our moving double pendulum in motion.
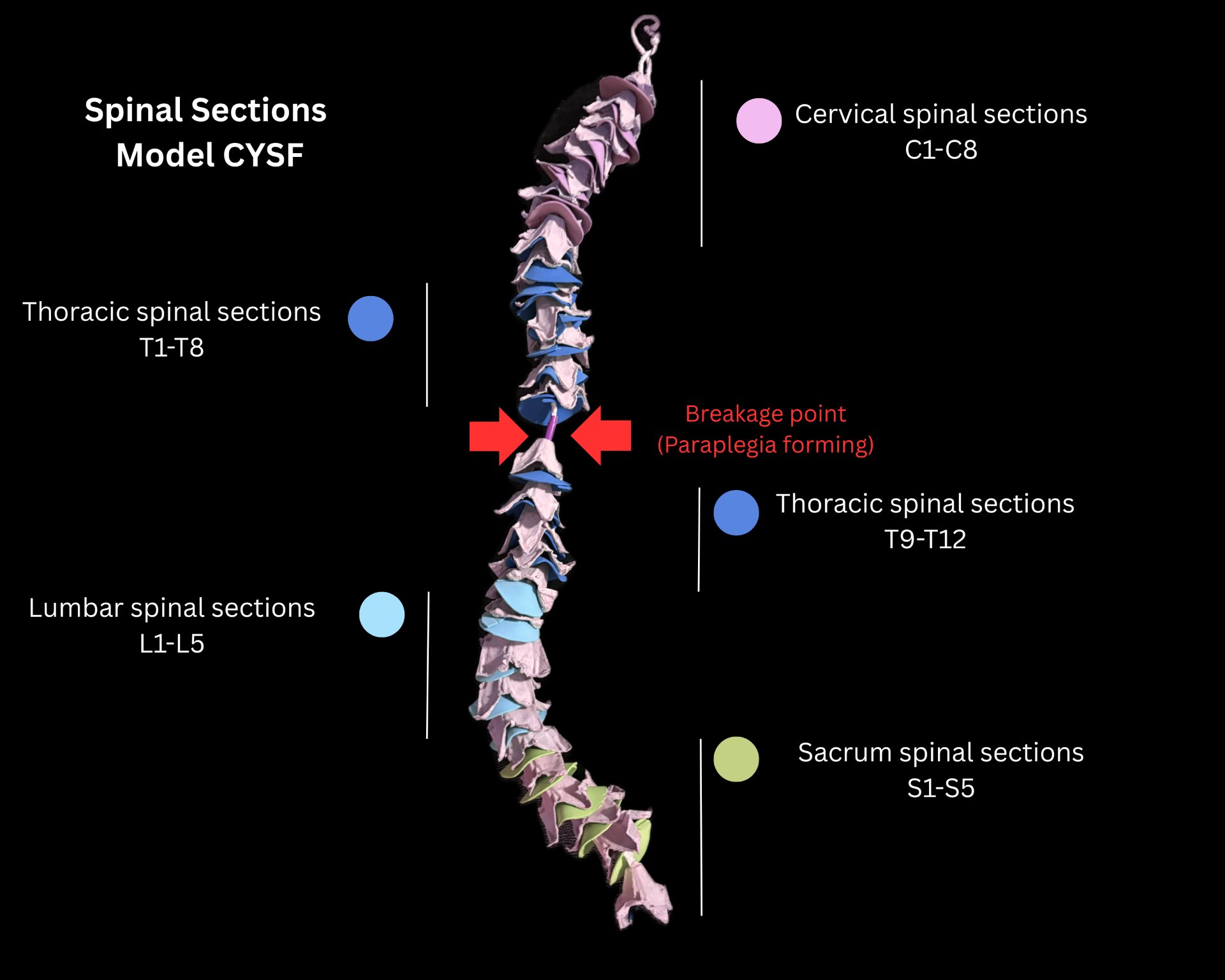
2.8.2 Physical Model (Spinal Cord Model)
To better understand the anatomy of the spinal cord with paraplegia, we created a rudimentary model.
2.9: Modelling Summary
In summary, this pendulum-based model that is going to be created will serve as a conceptual illustration of the principles behind dynamic scaffolds, and it connects the broader idea of dancing molecules in biomimetic materials and not fully relying on computational simulation. Also, creating the spinal cord makes it easier to understand what we are doing.
Analysis
3.1: Overview
The purpose of this analysis was to determine whether internal dynamic motion improves interaction frequency, alignment behaviour, and effective connectivity within a scaffold analogue system. Using a pendulum-based physical model, rigid pendulums represented static molecular structures, while double-jointed oscillatory pendulums represented dynamic "dancing molecules." Multiple controlled trials of equal duration were conducted to evaluate reproductivity and behavioural trends. Across repeated observations, dynamic systems consistently demonstrated enhanced interaction efficiency and adaptive alignment compared to static systems.
| Metric | Rigid Molecules | Dancing Molecules | Notes |
|---|---|---|---|
| Interaction Frequency | Low | High | Expected trend based on motion dynamics. Rigid swings are observed on one plane; dancing swings are multi-directional. |
| Alignment Recovery | Poor | Self-correcting | Oscillatory motion increases adaptive reorientation. A dynamic pendulum realigns towards the centre area after being pushed. |
| Receptor Coverage | Limited | Broad | Dynamic motion allows more receptor contact within the period of 10 s. |
| Probabilistic Efficiency | Low | High | Continuous motion increases the likelihood of successful interactions. |
| Visual Complexity | Simple | Complex | More dynamic motion is visible in oscillation patterns. |
Over the period of 10 s, we had counted the oscillations on both pendulums. The data are shown below:
| Pendulum Type | Trial 1 | Trial 2 | Trial 3 | Mean (Avg) |
|---|---|---|---|---|
| Rigid | 17 | 15 | 13 | 15 |
| Dancing | 22 | 23 | 19 | 20.7 |
The dynamic pendulum demonstrated a substantially higher mean oscillation count (20.7) compared to the rigid pendulum (15), representing a 38% increase in oscillatory activity over the 10-second interval. This increase supports the hypothesis that internal dynamic motion enhances interaction probability as well as system responsiveness.
Despite the static, one is still able to swing more than 10 times within 10 seconds; its motion isn't as consistent and gets more limited after each swing. The table below further expresses that:
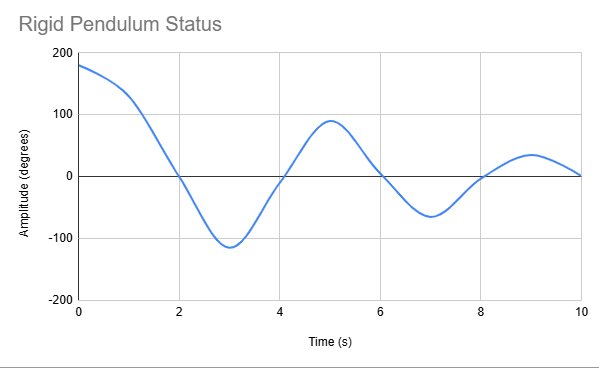
Rigid Pendulum Status:
| Time (s) | Amplitude | Note |
|---|---|---|
| 0 | 180 degrees | Initial pull |
| 5 | Reduced | Midpoint |
| 10 | Minimal | Near rest position |
Following our results, we constructed this table to further secure a clear understanding of simple pendulum motion.
On the other hand, the dynamic pendulum tends to maintain oscillation in a much more graceful way:
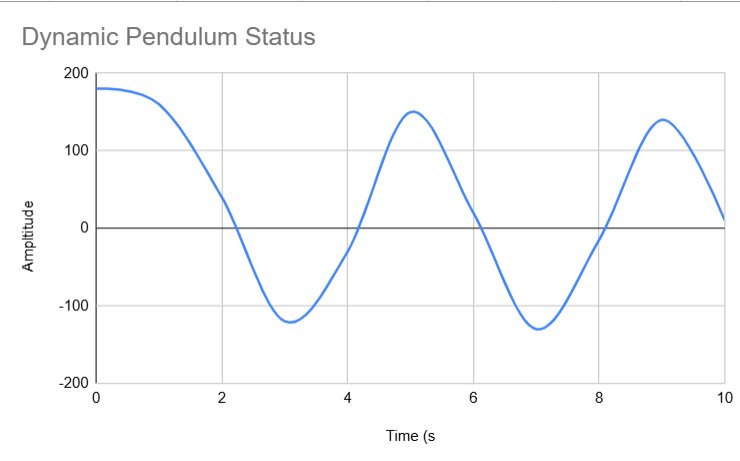
Dynamic Pendulum Status:
| Time (s) | Amplitude (rad) | Notes |
|---|---|---|
| 0 | 180 degrees | Initial/controlled point. |
| 5 | Sustained oscillation | Multi-axis motion is present. |
| 10 | Maintained Oscillation | Continuous adaptive movement. |
To mark its stronger forces of oscillation, we also created a graph for the double-jointed pendulum as well...
3.2: Interaction Frequency and Collision Probability
Dynamic pendulums exhibited a higher frequency of magnetic contact with the central spinal receptor zone than rigid pendulums. Their continuous oscillatory and lateral movement increased spatial exploration and surface coverage, leading to more frequent receptor engagement events within the same time interval. This finding illustrates a fundamental probabilistic principle: increased mobility increases collision likelihood. In biological systems, molecular binding and signal transduction depend on randomized collision events. By increasing amplitude and directional variability, dynamic elements effectively increase interaction probability without increasing forces. The consistency of this trend across trials indicated that motion-enabled systems systematically outperform static systems in generating functional contact events.
3.3: Alignment and Structural Reorientation
Following displacement or perturbation, dynamic pendulums demonstrated partial self-correcting alignment toward the receptor zone through continued oscillation. Over repeated cycles, they naturally reoriented toward equilibrium configurations. In contrast, rigid pendulums frequently remained misaligned after initial displacement, indicating limited adaptive capacity. This distinction is mechanically and biologically significant. In living tissues, extracellular matrix components such as collagen fibres reorganize along stress vectors to restore structural continuity. The dynamic model mirrors this adaptive behaviour, suggesting that mobility enhances resilience and recovery following mechanical disturbance. Thus, motion appears to function not only as an interaction amplifier but also as a structural stabilizer.
3.4: Signal Distribution and Spatial Coverage
In the model, magnets embedded in the receptor zone served as analogues for molecular binding sites or signal capture regions. Dynamic pendulums traversed a broader area of the receptor surface over time, whereas rigid pendulums contacted only limited, repetitive regions. This broader coverage indicates improved spatial distribution. From a transport perspective, increased motion reduces localized stagnation and increases uniformity of contact. Biologically, this behaviour is analogous to:
- Improved maintenance of chemical concentration gradients.
- Enhanced chemotactic guidance.
- Increased probability of electrical or biochemical signal capture.
Dynamic systems therefore demonstrate superior coverage efficiency, which may translate to improved signal dispersion in regenerative environments.
3.5: Reproducibility and Trend Consistency
Across multiple trials of equal duration, average interaction counts for dynamic pendulums consistently exceeded those of rigid molecules. Although minor variability occurred due to natural oscillatory differences, the directional trend remained stable. This reproducibility strengthens the internal validity of the findings and suggests that the observed advantage of motion-enabled systems is systematic rather than incidental. Future quantitative statistical testing could further confirm significance; however, even at the observational level, the trend is clear and repeatable.
3.6: Biological Relevance
The behaviour of dynamic pendulums parallels several regenerative biological processes, including axonal sprouting, extracellular matrix remodelling, and collagen fibre realignment under mechanical stress. In living systems, healing involves continuous structural reorganization rather than static replacement. The findings support a broader conceptual principle: effective biomimetic scaffolds must replicate not only structural architecture but also dynamic behaviour. Mobility appears to enhance interaction probability, alignment recovery, and spatial distribution—all of which are critical factors in tissue regeneration.
3.7: Limitations and Critical Evaluation
While the model demonstrates clear advantages of dynamic motion, several limitations must be acknowledged to contextualize the findings.
3.7.1: Scale and Physical Representation
The pendulum system is a macroscopic mechanical analogy rather than a molecular-scale replication. Molecular motion in biological tissues occurs at nanometer scales and is influenced by Brownian motion, thermal energy, and fluid dynamics. The present model simplifies these forces into gravitational oscillation and mechanical freedom. Therefore, while probabilistic interaction principles remain valid, the system does not replicate molecular biomechanics precisely.
3.7.2: Diffusion Not Directly Quantified
Improved signal dispersion was inferred from increased spatial coverage and contact frequency rather than from direct measurement of diffusion coefficients. In biological systems, diffusion is influenced by viscosity, molecular size, and extracellular spacing. Future iterations could incorporate quantitative diffusion mapping to strengthen transport-based conclusions.
3.7.3: Biological Complexity
The model does not incorporate immune responses, glial scar formation, inhibitory signalling, or cellular heterogeneity. Spinal cord regeneration is influenced by numerous biochemical and cellular factors beyond structural alignment. As such, the findings isolate mechanical mobility as a single variable rather than modelling the full regenerative cascade.
3.8: Balanced Interpretation
Despite these limitations, the findings consistently demonstrate that internal dynamic motion enhances interaction frequency, spatial coverage, and alignment recovery within the scaffold analogue system. The model does not claim to replicate complete biological regeneration. Instead, it tests a foundational principle: that movement itself may function as a regenerative enhancer. By identifying motion as an active design parameter rather than a passive property, this study suggests a potential shift in scaffold engineering philosophy from static, structural mimicry toward dynamic, behavioural mimicry.
3.8.1: Therapeutic Applications
Based on real-world scientific evidence, especially animal studies with paralyzed mice, a single injection of the "dancing molecules" therapy led to significant functional improvement, which allowed the mice to regain the ability to walk within four weeks. The treatment achieves this by:
- Regenerative axons
- Significantly diminishing glial scar tissue (Physical barrier to regeneration)
- Reforming Myelin (insulating layer around axons)
- Forming new/functional blood vessels
- More efficient motor neurons
All this requires specialized materials that are designed to be temporary and would biodegrade into nutrients for the cells within \~12 weeks; then they would disappear completely from the body without noticeable side effects. Furthermore, the therapy has also shown promise in lab-grown human spinal cord organoids.
3.8.2: Current Status
For advancing innovations like this, it is really important to know where the path to such revolutionizing concepts is heading. Researchers, primarily from Northwestern University, are currently collecting data to seek FDA approval to begin human clinical trials. The underlying principle of "supramolecular motion" as a key factor in bioactivity is also being explored for other regenerative medicine applications, including for cartilage regeneration for conditions like osteoarthritis and for treating neurodegenerative diseases such as Alzheimer's, Parkinson's, and stroke.
Conclusion
This project demonstrates that internal dynamic motion—exemplified by the "dancing molecules" concept—can significantly enhance interaction frequency, alignment recovery, and structural adaptability within biomimetic scaffold models. Through both computational and physical pendulum-based simulations, dynamic pendulum consistency exhibited greater motion coverage, higher collision probabilities, and improved self-correcting behaviour compared to rigid analogues. These findings highlight the critical role of mobility as an active facilitator of molecular contact and regenerative potential, rather than a passive structural property.
The observed trends align with biological phenomena such as axonal sprouting, extracellular matrix remodelling, and collagen fibre realignment, suggesting that motion-responsive scaffolds more accurately replicate the behavioural characteristics of living tissues. For conditions like paraplegia, where spinal cord injuries disrupt limb function, these dynamic scaffolds may provide improved guidance for axonal regrowth, promoting recovery of motor and sensory function.
While this project relied on simplified models and simulated data, it provides foundational evidence that motion-enhanced designs offer clear advantages over static scaffolds. These insights show the importance of behavioural mimicry in regenerative medicine and suggest developing advanced, adaptive scaffolds capable of supporting tissue healing, neural repair, and improved outcomes for patients with spinal cord injuries.
In summary, integrating dynamic motion into biomimetic scaffold design represents a strong shift—from static architectural replication toward functional, adaptive regeneration—and establishes a compelling framework for future research and clinical translation.
Citations
Engelhardt, B. (2006). Molecular mechanisms involved in T cell migration across the blood–brain barrier. Journal of Neural Transmission, 113(4), 477–485.
Li, W., Liu, G., Liang, J., Wang, X., Song, M., Liu, X., Wang, L., Yang, Z., & Zhang, B. (2025). The dance between Schwann cells and macrophages during the repair of peripheral nerve injury. Neuroscience Bulletin, 41(8), 1448–1462.
Matsuzaka, Y., & Yashiro, R. (2023). Unraveling the immunopathogenesis of multiple sclerosis: The dynamic dance of plasmablasts and pathogenic T cells. Biologics, 3(3), 232–252.
Naeem, A., Yu, C., Zhou, L., Xie, Y., Weng, Y., Huang, Y., Zhang, M., & Yang, Q. (2025). Shape memory hydrogels in tissue engineering: Recent advances and challenges. Bioactive Materials, 54, 215–247.
Pfau, M. R., & Grunlan, M. A. (2021). Smart scaffolds: Shape memory polymers (SMPs) in tissue engineering. Journal of Materials Chemistry B, 9(21), 4287–4297.
Stupp, S. I. (2021, November 11). ‘Dancing molecules’ successfully repair severe spinal cord injuries. Northwestern Now. Weintraub, A. (2021, November 15).
Weintraub, A. (2021). Injectable'dancing molecules' repair spinal cord injury, restore mobility in paralyzed mice. Fiercebiotech. com, NA-NA.
Balint, R., Cassidy, N. J., & Cartmell, S. H. (2014). Conductive polymers: Towards a smart biomaterial for tissue engineering. Acta Biomaterialia, 10(6), 2341–2353.
Discher, D. E., Janmey, P., & Wang, Y. (2005). Tissue cells feel and respond to the stiffness of their substrate. Science, 310(5751), 1139–1143.
Frantz, C., Stewart, K. M., & Weaver, V. M. (2010). The extracellular matrix at a glance. Journal of Cell Science, 123(24), 4195–4200.
Fratzl, P. (2008). Collagen: Structure and mechanics. Springer.
Hollister, S. J. (2005). Porous scaffold design for tissue engineering. Nature Materials, 4(7), 518–524.
Ingber, D. E. (2006). Cellular mechanotransduction: Putting all the pieces together again. The FASEB Journal, 20(7), 811–827.
Silver, J., & Miller, J. H. (2004). Regeneration beyond the glial scar. Nature Reviews Neuroscience, 5(2), 146–156.
Chen, Z., Wang, L., Chen, C., Sun, J., Luo, J., Cui, W., … & Shi, Q. (2022). NSC‑derived extracellular matrix‑modified GelMA hydrogel fibrous scaffolds promote neural stem cell adhesion and differentiation and improve spinal cord repair. NPG Asia Materials.
Zheng, Y., Nützl, M., Schackel, T., Chen, J., Weidner, N., Müller, R., & Puttagunta, R. (2024). Biomaterial scaffold stiffness influences neuroregeneration in spinal cord injury. Bioactive Materials.
Slotkin, J. R., et al. (2017). Biodegradable scaffolds promote tissue remodeling and functional improvement in non‑human primates with acute spinal cord injury. Biomaterials.
Mangani, S., Vetoulas, M., Mineschou, K., et al. (2025). Design and applications of extracellular matrix scaffolds in tissue engineering and regeneration. Cells.
Neural Regeneration Research (2025). Stiffness‑tunable biomaterials provide a good environment for neural growth and regeneration. Neural Regeneration Research.
Karki, K. T. (2024). Dancing Molecules. Eastern Green Neurosurgery, 5(01), 1-2.
‘Dancing molecules’ successfully repair severe spinal cord injuries - Northwestern Now.
Alberts, A., Lungescu, I. A., Niculescu, A.-G., & Grumezescu, A. M. (2025). Natural Products for Improving Soft Tissue Healing: Mechanisms, Innovations, and Clinical Potential. Pharmaceutics, 17(6), 758
Ju, W., Santos, A., Freeman, A., & Daniele, E. (2018). Neuroscience: Canadian 1st Edition [Open textbook]. eCampusOntario.
https://byjus.com/biology/spinal-cord/
Kasukawa, Y., Shimada, Y., Kudo, D., Saito, K., Kimura, R., Chida, S., Hatakeyama, K., & Miyakoshi, N. (2022). Advanced Equipment Development and Clinical Application in Neurorehabilitation for Spinal Cord Injury: Historical Perspectives and Future Directions. Applied Sciences, 12(9), 4532.
Onorato, P., & De Ambrosis, A. (2012). Magnetic damping: Integrating experimental and theoretical analysis. American journal of physics, 80(1), 27-35.
Crossley, R. M., Johnson, S., Tsingos, E., Bell, Z., Berardi, M., Botticelli, M., ... & Shuttleworth, R. (2024). Modeling the extracellular matrix in cell migration and morphogenesis: a guide for the curious biologist. Frontiers in cell and developmental biology, 12, 1354132.
Dixon, D. T., Landree, E. N., & Gomillion, C. T. (2025). Biomimetic dual‑sensing bone scaffolds: Characterization and in vitro evaluation under dynamic culturing conditions. Biomimetics, 10(9), 598.
Liu, J., Wang, X., Wu, Z., Hui, Z., Zhang, J., Zhang, J., … Yang, X. (2026). Dynamic stimulation of piezoelectric scaffolds enhances osteogenesis‑related biological responses via electro‑mechanical sensitive channels and cytoskeletal remodeling. Biomaterials, 329, 123938.
Fan, S., Qian, Y., Li, D., Lu, L., Lian, J., Liu, X., … Yang, L. (2025). Development of a user‑friendly dynamic culture system for decellularized scaffold‑based tissue engineering. ACS Biomaterials Science & Engineering, 11(10), 6272–6281.
Karki, K. T. (2024). Dancing Molecules. Eastern Green Neurosurgery, 5(01), 1-2.
Fulcher, L. P., & Davis, B. F. (1976). Theoretical and experimental study of the motion of the simple pendulum. American Journal of Physics, 44(1), 51-55.
Calvão, A. M., & Penna, T. J. P. (2015). The double pendulum: a numerical study. European Journal of Physics, 36(4), 045018.
Levien, R. B., & Tan, S. M. (1993). Double pendulum: An experiment in chaos. American Journal of Physics, 61, 1038-1038.
Herrera-Arozamena, C., Martí-Marí, O., Estrada, M., De la Fuente Revenga, M., & Rodríguez-Franco, M. I. (2016). Recent advances in neurogenic small molecules as innovative treatments for neurodegenerative diseases. Molecules, 21(9), 1165.
Kazim, S. F., & Iqbal, K. (2016). Neurotrophic factor small-molecule mimetics mediated neuroregeneration and synaptic repair: emerging therapeutic modality for Alzheimer’s disease. Molecular neurodegeneration, 11(1), 50.
Bromley, I. (2006). Tetraplegia and paraplegia: a guide for physiotherapists. Elsevier Health Sciences.
Figoni, S. F. (1993). Exercise responses and quadriplegia. Medicine and science in sports and exercise, 25(4), 433-441.
Álvarez, Z., Kolberg-Edelbrock, A. N., Sasselli, I. R., Ortega, J. A., Qiu, R., Syrgiannis, Z., ... & Stupp, S. I. (2021). Bioactive scaffolds with enhanced supramolecular motion promote recovery from spinal cord injury. Science, 374(6569), 848-856.
Du, G., Moulin, E., Jouault, N., Buhler, E., & Giuseppone, N. (2012). Muscle‐like supramolecular polymers: integrated motion from thousands of molecular machines. Angewandte Chemie, 124(50), 12672-12676.
Kim, H. J., Lee, E., Park, H. S., & Lee, M. (2007). Dynamic extension− contraction motion in supramolecular springs. Journal of the American Chemical Society, 129(36), 10994-10995.
Cheng, M., Zhu, G., Li, L., Zhang, S., Zhang, D., Kuehne, A. J., & Shi, F. (2018). Parallel and precise macroscopic supramolecular assembly through prolonged Marangoni motion. Angewandte Chemie, 130(43), 14302-14306.
Acknowledgement
We would like to express our sincere gratitude to our science fair coordinators Ms. Friesen & Dr. Zia for their guidance, thoughtful feedback, and encouragement throughout the development of this project–as their support helped refine both the scientific depth and clarity of the overall project.
We are especially thankful for the opportunity to explore interdisciplinary concepts that connect biology, physics, and materials science. This project would not have been possible without access to educational resources, scientific literature, and constructive academic discussions that are strengthened by understanding of regenerative medicine and biomimetic design.
We would also like to acknowledge the pioneering researchers in the field of supramolecular chemistry and regenerative biomaterials whose published work inspired the foundational ideas behind this study. Their discoveries in motion-responsive molecular systems provided the conceptual framework for exploring the “Dancing Molecules Effect” in a simplified mechanical model.
We are also grateful to our families, who have always been supportive of our efforts, and who helped to motivate us to explore these ambitious scientific ideas.
Finally, we are grateful for the opportunity in this science fair, which further encouraged our curiosity, creativity, and innovation among young researchers.